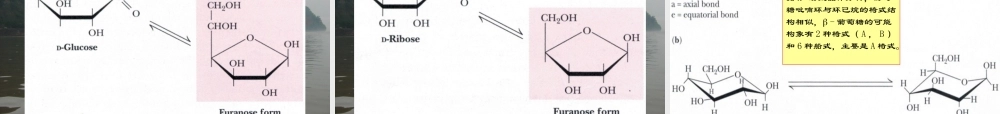
第1章糖类一、引言(一)糖类的存在与来源碳水化合物是地球上最丰富的有机化合物,每年全球植物和藻类光合作用可转换1000亿吨CO2和H2O成为纤维素和其他植物产物。植物体85-90%的干重是糖类。总的说来,糖类在生物体内所起的作用包括:能量物质、结构物质和活性物质。(二)糖类的生物学作用绿色植物的皮、杆等的多糖(纤维素、半纤维素和果胶物质等);昆虫、蟹、虾等外骨骼糖(几丁质);结缔组织中的糖(肝素、透明质酸、硫酸软骨素、硫酸皮肤素等);细菌细胞壁糖称作结构多糖;粮食及块根、块茎中的多糖(淀粉);动物体内的贮藏多糖(糖元)是重要的能源物质;糖蛋白(蛋白聚糖)中的糖;细胞膜及其他细胞结构中的糖如血型糖;活性糖分子是重要的信息分子;医疗用糖(葡萄糖及其衍生物,如葡萄糖酸的钠、钾、钙、锌盐等);食用菌中的糖(香菇多糖、茯苓多糖、灵芝多糖、昆布多糖等)可以作为药物使用;糖类是重要的中间代谢物,可以转化为氨基酸、核苷酸和脂类。糖类主要由C、H、O三种元素组成,有些还有N、S、P等。单糖多符合结构通式:(CH2O)n,但仅从通式上并不能判断某分子是否就是糖,即:符合通式的不一定是糖,如CH3COOH(乙酸),CH2O(甲醛),C3H6O3(乳酸);是糖的不一定都符合通式,如C5H10O4(脱氧核糖),C6H12O5(鼠李糖)。糖类可以定义为:多羟基醛;多羟基酮;多羟基醛或多羟基酮的衍生物;可以水解为多羟基醛或多羟基酮或它们的衍生物的物质。(三)糖类的元素组成和化学本质(四)糖的命名与分类单糖(monosaccharides)不能水解为其他糖的糖,按碳原子数分为:丙糖(甘油醛);丁糖(赤藓糖);戊糖(木酮糖、核酮糖、核糖、脱氧核糖等);己糖(葡萄糖、果糖、半乳糖等)等。寡糖(oligosaccharides)可以水解为几个至十几个单糖的糖,一般包括:二糖(disaccharides)蔗糖、麦芽糖、乳糖;三糖(trisaccharides)棉籽糖和其他寡糖。多糖(polysaccharides)可水解为多个单糖或其衍生物的糖,包括:同多糖(homoglycans,homopolysaccharides)水解为同一单糖的高分子聚合物,淀粉、糖元、纤维素、几丁质、糖苷等。异多糖(heteroglycans,heteropolysaccharides)水解产物不止一种单糖或单糖衍生物,透明质酸、肝素、硫酸软骨素、硫酸皮肤素等。糖的衍生物指糖的氧化产物、还原产物、氨基取代物及糖苷化合物等,如,D-氨基葡萄糖、N-乙酰氨基葡萄糖、糖的硫酸酯等;多糖复合物(polysaccharidescomplex)糖与脂、蛋白...