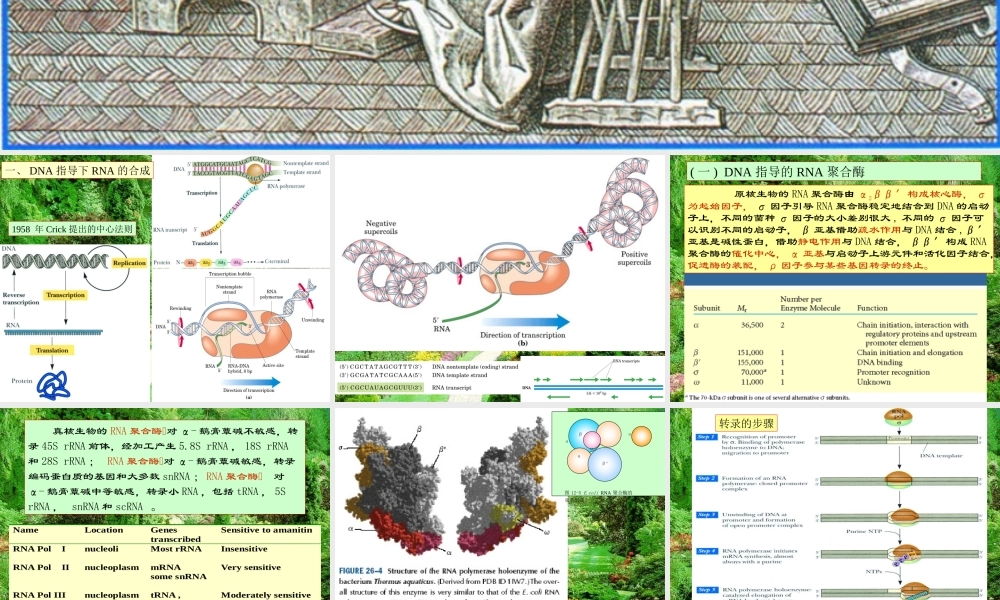
生物合成及其调控第36章RNA的一、DNA指导下RNA的合成1958年Crick提出的中心法则(一)DNA指导的RNA聚合酶原核生物的RNA聚合酶由α2ββ′构成核心酶,σ为起始因子,σ因子引导RNA聚合酶稳定地结合到DNA的启动子上,不同的菌种σ因子的大小差别很大,不同的σ因子可以识别不同的启动子,β亚基借助疏水作用与DNA结合,β′亚基是碱性蛋白,借助静电作用与DNA结合,ββ′构成RNA聚合酶的催化中心,α亚基与启动子上游元件和活化因子结合,促进酶的装配,ρ因子参与某些基因转录的终止。NameLocationGenestranscribedSensitivetoamanitinRNAPolInucleoliMostrRNAInsensitiveRNAPolIInucleoplasmmRNAsomesnRNAVerysensitiveRNAPolIIInucleoplasmtRNA,5SrRNAU6snRNA,smallRNAModeratelysensitive真核生物的RNA聚合酶对α-鹅膏蕈碱不敏感,转录45SrRNA前体,经加工产生5.8SrRNA,18SrRNA和28SrRNA;RNA聚合酶对α-鹅膏蕈碱敏感,转录编码蛋白质的基因和大多数snRNA;RNA聚合酶对α-鹅膏蕈碱中等敏感,转录小RNA,包括tRNA,5SrRNA,snRNA和scRNA。βασωαβ’图12-5E.coliRNA聚合酶的亚基组成转录的步骤研究转录起点的方法:足迹法(二)启动子和转录因子原核生物的启动子不同的σ因子识别的启动子共有序列有所不同(表36-3)真核生物RNA聚合酶启动子的共有序列RNA聚合酶基本启动子的共有序列TATA位于-25至-30,转录起始部位有一保守序列PyPyA*NT(A)PyPy,其中的Py表示嘧啶,*表示+1位点。上游调控元件包括CAAT框,GC框和八聚体框等,位置不很确定,不同的细胞可以有不同的上游调控元件,不同的上游调控元件与不同的转录因子相互作用(表36-4)。真核生物RNA聚合酶启动子与转录因子的相互作用真核生物RNA聚合酶前转录复合物TFⅡD是包含TATA结合蛋白(TATA-bindingprotein,TBP)和多种TBP联结因子(TBP-associatedfactor,TAF)的寡聚蛋白,可以与RNA聚合酶Ⅱ的C端相互作用。TFⅡB有两个结构域,一个结合TBP。另一个可以引进TFⅡF/polⅡ复合物TFⅡF有ATP酶,解螺旋酶,激酶等多种活性,可以使RNA聚合酶Ⅱ大亚基的C端磷酸化,引起构像变化,促进转录。还可以参与DNA损伤的修复。ⅡE和ⅡH促进除ⅡF以外的其他转录因子脱落,使转录由起始阶段进入延伸阶段。黄色为TATAbox的糖磷酸骨架,碱基对为红色,相邻的DNA片段为蓝色,马鞍形的TBP(绿色)结合在DNA的小沟,使小沟扩大,并使DNA轴弯曲约100o,使TATA序列解旋。TFⅡD杂聚体的其它组分位于TBP上方,...