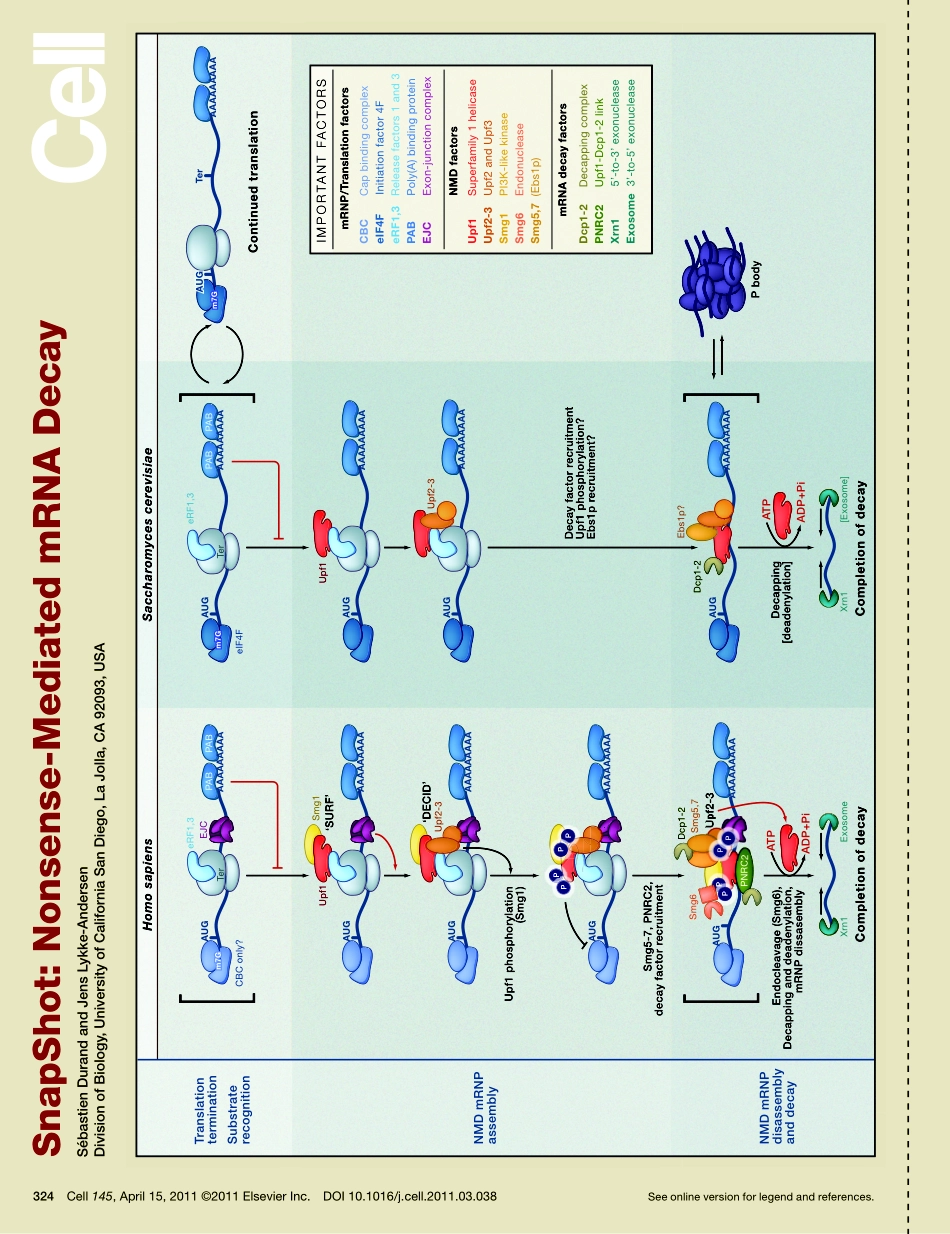
SnapShot:Nonsense-MediatedmRNADecaySébastienDurandandJensLykke-AndersenDivisionofBiology,UniversityofCaliforniaSanDiego,LaJolla,CA92093,USASeeonlineversionforlegendandreferences.324Cell145,April15,2011©2011ElsevierInc.DOI10.1016/j.cell.2011.03.038PPPPPPPm7Gm7Gm7Gm7Gm7Gm7GAUGAUGAUGm7Gm7Gm7GTranslationterminationSubstraterecognitionNMDmRNPassemblyIMPORTANTFACTORSmRNP/TranslationfactorsCBCCapbindingcomplexeIF4FInitiationfactor4FeRF1,3Releasefactors1and3PABPoly(A)bindingproteinEJCExon-junctioncomplexUpf1Superfamily1helicaseUpf2-3Upf2andUpf3Smg1PI3K-likekinaseSmg6EndonucleaseSmg5,7(Ebs1p)DecayfactorrecruitmentUpf1phosphorylation?Ebs1precruitment?Smg5-7,PNRC2,decayfactorrecruitmentPNRC2Dcp1-2Dcp1-2Upf2-3Upf2-3Smg5,7Ebs1p?Smg6Xrn1ExosomeXrn1[Exosome]CBConly?eIF4FTerTerEJCUpf1Upf1Smg1PABPABPABPABeRF1,3eRF1,3Upf2-3ATPADP+PiATPADP+PiUpf1phosphorylation(Smg1)‘SURF’‘DECID’AAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAUGAUGAUGAUGAUGAUGAUGAUGTerAUGAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAAEndocleavage(Smg6),Decappinganddeadenylation,mRNPdissasemblyDecapping[deadenylation]PbodyCompletionofdecayCompletionofdecayContinuedtranslationDcp1-2DecappingcomplexPNRC2Upf1-Dcp1-2linkXrn15’-to-3’exonucleaseExosome3’-to-5’exonucleaseNMDfactorsmRNAdecayfactorsNMDmRNPdisassemblyanddecaySaccharomycescerevisiaeHomosapiensSnapShot:Nonsense-MediatedmRNADecaySébastienDurandandJensLykke-AndersenDivisionofBiology,UniversityofCaliforniaSanDiego,LaJolla,CA92093,USA324.e1Cell145,April15,2011©2011ElsevierInc.DOI10.1016/j.cell.2011.03.038Thenonsense-mediatedmRNAdecay(NMD)pathwayservesanimportantfunctioninmRNAqualitycontrolbyriddingthecellofaberrantmRNAsthatencodetruncatedproteinsduetoprematuretranslationterminationcodons(PTCs).PTC-containingmRNAsarisefromerrorsintranscriptionorpre-mRNAprocessingorfrommutationorrecombinationeventswithinprotein-codinggenes.Theyarealsoproducedfromalternativepre-mRNAsplicingevents.Inaddition,asubsetofnormallyexpressedmRNAsaredownregulatedbytheNMDpathway(reviewe...