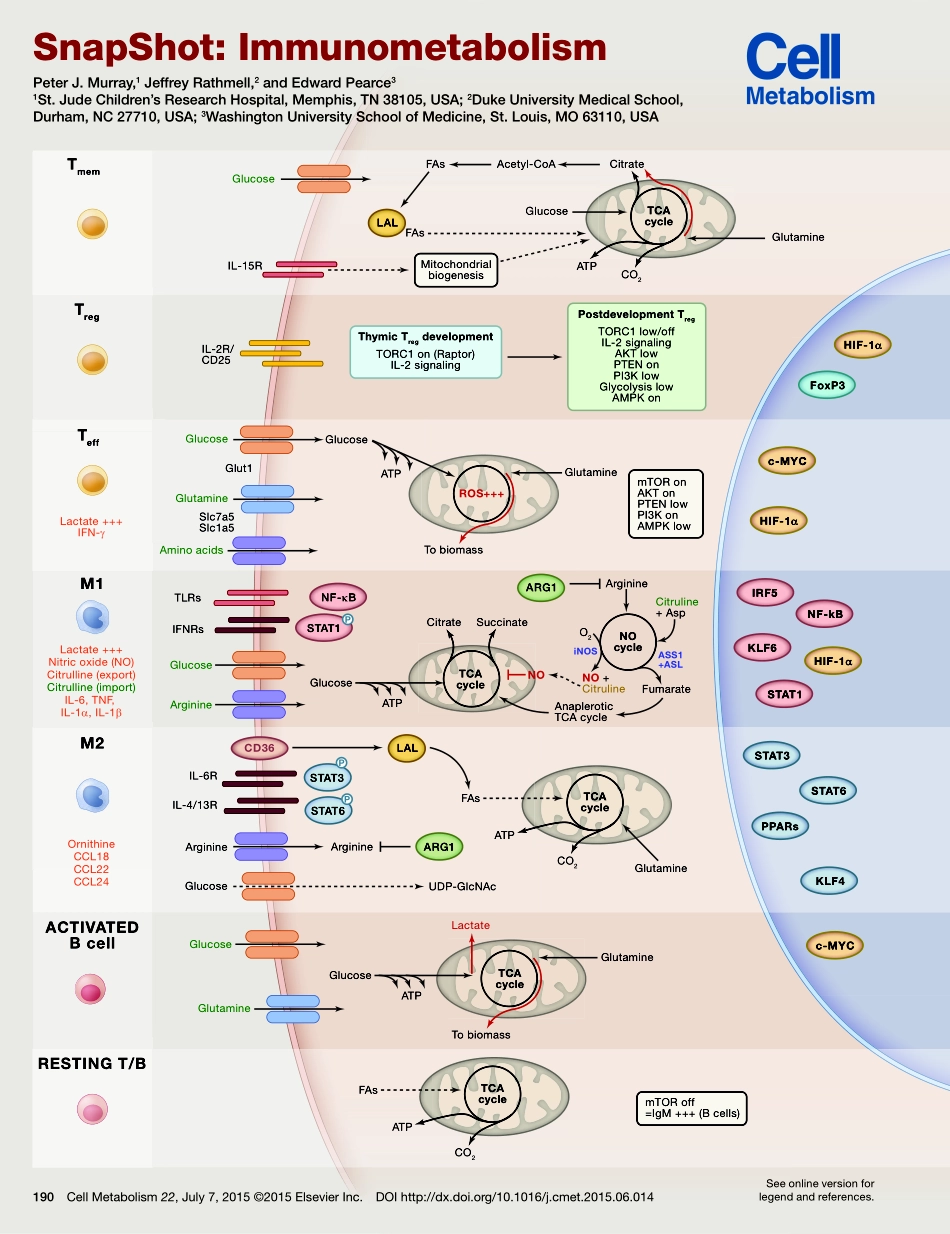
Lactate+++Nitricoxide(NO)Citrulline(export)Citrulline(import)IL-6,TNF,IL-1α,IL-1βOrnithineCCL18CCL22CCL24Lactate+++IFN-γSlc7a5Slc1a5Glut1FAsFAsMitochondrialbiogenesisThymicTregdevelopmentTORC1on(Raptor)IL-2signalingPostdevelopmentTregTORC1low/offIL-2signalingAKTlowPTENonPI3KlowGlycolysislowAMPKonmTORonAKTonPTENlowPI3KonAMPKlowmTORoff=IgM+++(Bcells)Acetyl-CoACitrateGlucoseGlucoseTLRsIFNRsIL-6RIL-4/13RGlucoseGlucoseLactateGlucoseGlutamineGlutamineAminoacidsArginineIL-15RIL-2R/CD25GlucoseGlutamineTobiomassO2iNOSASS1+ASLArginineNO+CitrulineCitruline+AspFumarateAnapleroticTCAcycleGlutamineM2M1TeffTregTmemACTIVATEDBcellRESTINGT/BHIF-1αHIF-1αHIF-1αFoxP3c-MYCc-MYCIRF5NF-kBNF-κBKLF6STAT1STAT1STAT3STAT6STAT6ARG1ARG1STAT3CD36PPARsKLF4LALLALGlucoseGlucoseArginineArginineUDP-GlcNAcTCAcycleNOcycleROS+++GlutamineFAsCO2ATPTCAcycleCO2ATPCitrateSuccinateFAsCO2ATPTCAcycleTCAcycleATPTobiomassGlutamineGlucoseATPATPPPPNOOrnithineCCL18CCL22M2Lactate+++Nitricoxide(NO)Citrulline(export)Citrulline(import)AminoacidsM1ACTIVATEDBcellRESTINGT/BRESTINGT/BTmemTregLactate+++IFN-γTeffAminoacidsTTCAcycleSnapShot:ImmunometabolismPeterJ.Murray,1JeffreyRathmell,2andEdwardPearce31St.JudeChildren’sResearchHospital,Memphis,TN38105,USA;2DukeUniversityMedicalSchool,Durham,NC27710,USA;3WashingtonUniversitySchoolofMedicine,St.Louis,MO63110,USASeeonlineversionforlegendandreferences.190CellMetabolism22,July7,2015©2015ElsevierInc.DOIhttp://dx.doi.org/10.1016/j.cmet.2015.06.014190.e1CellMetabolism22,July7,2015©2015ElsevierInc.DOIhttp://dx.doi.org/10.1016/j.cmet.2015.06.014SnapShot:ImmunometabolismPeterJ.Murray,1JeffreyRathmell,2andEdwardPearce31St.JudeChildren’sResearchHospital,Memphis,TN38105,USA2DukeUniversityMedicalSchool,Durham,NC27710,USA3WashingtonUniversitySchoolofMedicine,St.Louis,MO63110,USAThevastandfast-movingfieldofimmunometabolismdeciphersthecontributionofbiochemistrytoimmunecelldevelopment,fate,andbehavior.Anadvantageofusingtheimmunesystemtouncoverfundamentalaspectsofintermediarymetabo...