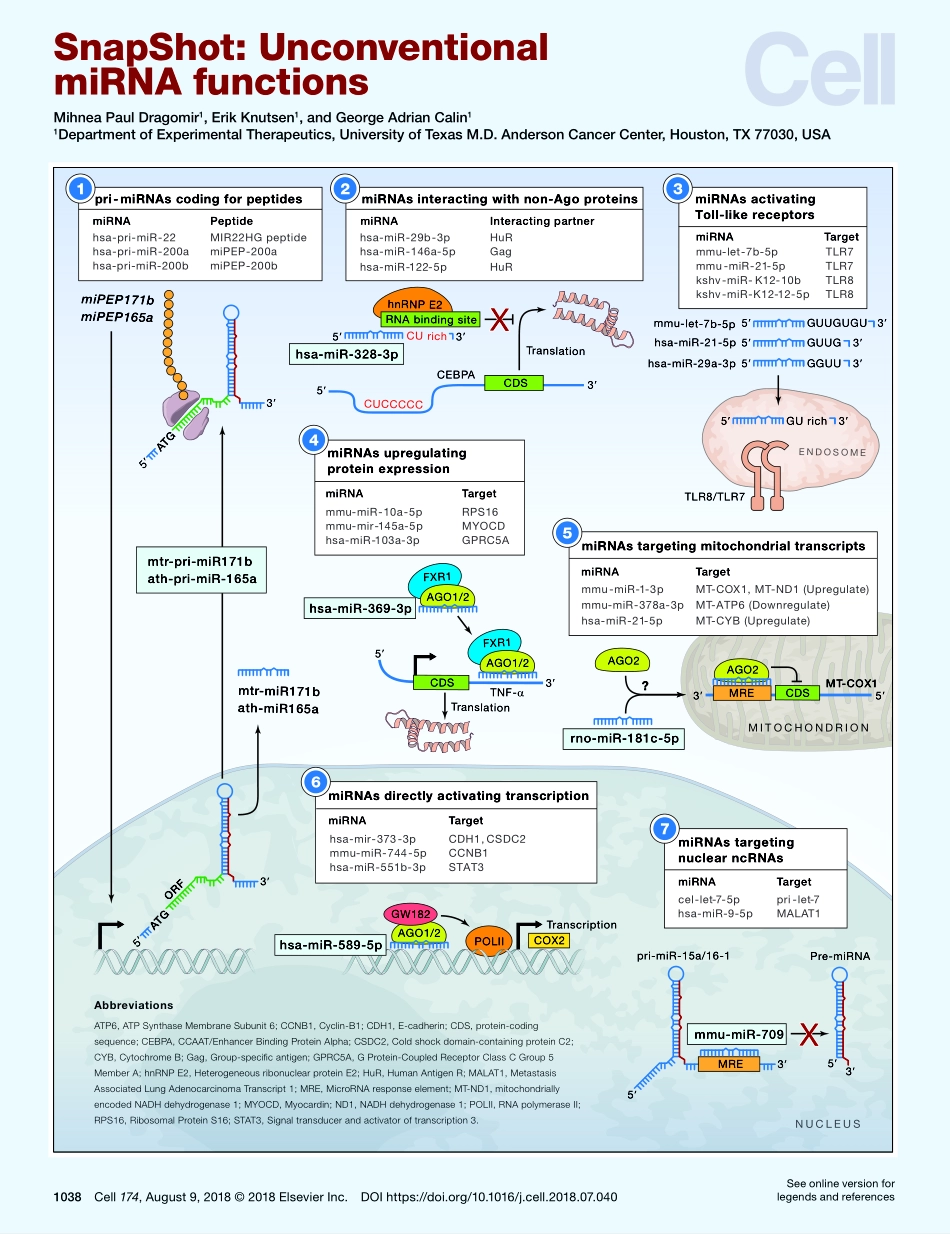
miPEP171bmiPEP165aMITOCHONDRIONmtr-miR171bath-miR165aORFATG35ATG35mtr-pri-miR171bath-pri-miR-165apri-miRNAscodingforpeptideshsa-pri-miR-22miRNAPeptideMIR22HGpeptidehsa-pri-miR-200ahsa-pri-miR-200bmiPEP-200amiPEP-200bmiRNAstargetingmitochondrialtranscriptsmmu-miR-1-3pMT-COX1,MT-ND1(Upregulate)mmu-miR-378a-3pMT-ATP6(Downregulate)hsa-miR-21-5pMT-CYB(Upregulate)MREAGO2MT-COX1CDS5315TLR8/TLR7ENDOSOMEGUUGUGUmmu-let-7b-5pGUUGGGUUhsa-miR-29a-3phsa-miR-21-5p353355GUrich35miRNAsactivatingToll-likereceptorsmmu-let-7b-5pTLR7mmu-miR-21-5pTLR7kshv-miR-K12-10bkshv-miR-K12-12-5pTLR8TLR83miRNATargetPOLIIAGO1/2GW182TranscriptionCOX2hsa-miR-589-5pmiRNAsdirectlyactivatingtranscriptionhsa-mir-373-3pCDH1,CSDC2mmu-miR-744-5pCCNB1hsa-miR-551b-3pSTAT36miRNATargetPre-miRNApri-miR-15a/16-13535MREmmu-miR-709XmiRNAstargetingnuclearncRNAscelhsa-miR-9-5pMALAT1-let-7-5ppri-let-77miRNATargetmiRNATargetTranslation335CUrichCUCCCCChnRNPE2RNAbindingsiteCDSXhsa-miR-29b-3pHuRhsa-miR-146a-5pGaghsa-miR-122-5pHuRmiRNAsinteractingwithnon-Agoproteins52CEBPAmiRNAInteractingpartnerhsa-miR-328-3pAGO2?rno-miR-181c-5pNUCLEUSTNF-αTranslationFXR1AGO1/2FXR1AGO1/2CDSmiRNAsupregulatingproteinexpressionmmu-miR-10a-5pRPS16mmu-mir-145a-5pMYOCDhsa-miR-103a-3pGPRC5A354miRNATargethsa-miR-369-3pATP6,ATPSynthaseMembraneSubunit6;CCNB1,Cyclin-B1;CDH1,E-cadherin;CDS,protein-codingsequence;CEBPA,CCAAT/EnhancerBindingProteinAlpha;CSDC2,Coldshockdomain-containingproteinC2;CYB,CytochromeB;Gag,Group-specificantigen;GPRC5A,GProtein-CoupledReceptorClassCGroup5MemberA;hnRNPE2,HeterogeneousribonuclearproteinE2;HuR,HumanAntigenR;MALAT1,MetastasisAssociatedLungAdenocarcinomaTranscript1;MRE,MicroRNAresponseelement;MT-ND1,mitochondriallyencodedNADHdehydrogenase1;MYOCD,Myocardin;ND1,NADHdehydrogenase1;POLII,RNApolymeraseII;RPS16,RibosomalProteinS16;STAT3,Signaltransducerandactivatoroftranscription3.Abbreviations1038Cell174,August9,2018©2018ElsevierInc.DOIhttps://doi.org/10.1016/j.cell.2018.07.040SeeonlineversionforlegendsandreferencesSnap...