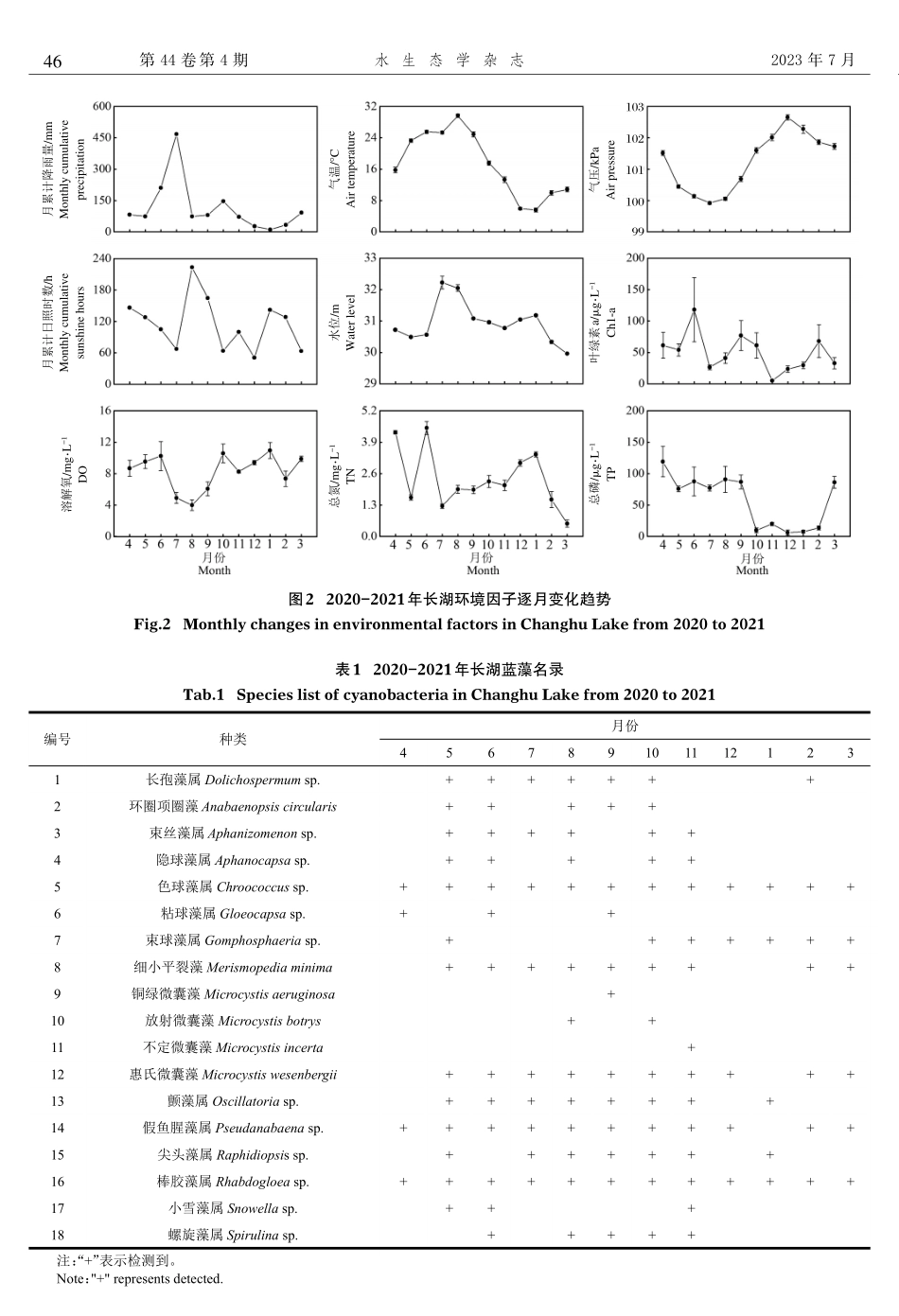
2023年7月水生态学杂志第44卷第4期收稿日期:2022-04-01修回日期:2022-09-30基金项目:国家自然科学基金(41901135;U21A2039);长江科学院开放基金(CKWV2019770/KY);湖北省教育厅项目(Q20191303)。作者简介:李滔,1998年生,男,硕士研究生,研究方向为水环境与水生态。E-mail:taolisy@163.com通信作者:杨军,1986年生,男,副教授,主要从事水环境与水生态、浮游植物和湿地水体修复研究。E-mail:junyang2@yangtzeu.edu.cnDOI:10.15928/j.1674-3075.202204010095湖北长湖蓝藻季节动态及其驱动因子分析李滔1,章叶飞1,王海艳1,熊勤学1,2,张露1,习小杰1,陈如1,刘章勇1,2,朱建强1,2,杨军1,2(1.长江大学农学院,湖北荆州434025;2.长江大学湿地生态与农业利用教育部工程研究中心,湖北荆州434025)摘要:长湖是调控长江中游地区生态平衡重要的湿地生态系统,为了解长湖蓝藻季节动态并进一步探究蓝藻与环境因子的相关关系,根据其湖形特征设置5个采样点,于2020年4月至2021年3月对长湖蓝藻及水环境进行了逐月采样调查与分析。结果显示,调查期间共检出蓝藻18种(属),主要优势种为棒胶藻属(Rhabdogloeasp.)、惠氏微囊藻(Microcystiswesenbergii)和假鱼腥藻属(Pseudanabaenasp.)。二维非度量多维尺度(NMDS)分析显示,长湖蓝藻生物量季节变化明显,5、6、8月的蓝藻生物量较高,为6.88~15.50mg/L,占浮游植物总生物量的24.0%~43.8%;冬季和初春较低,为0.13~0.29mg/L,占浮游植物总生物量的0.5%~2.8%。逐步回归分析显示,气温、气压、总磷、降雨量和溶解氧是影响蓝藻的重要因子,共解释了蓝藻生物量变化的70.0%。结构方程模型展示了环境因子与蓝藻生物量的关系,气温和溶解氧的上升可促进蓝藻生长,而气压、总磷和降雨量正好相反,环境因子可直接或间接地通过改变湖泊的湖沼学特征影响蓝藻生物量。研究结果可为长湖蓝藻水华防控提供参考依据。关键词:长湖;蓝藻;季节动态;环境因子中图分类号:Q948.11文献标志码:A文章编号:1674-3075(2023)04-0044-08水生态学杂志JournalofHydroecologyVol.44,No.4第44卷第4期2023年7月Jul.2023随着全球人口激增以及经济和社会的快速发展,大量的湖泊、河流和水库暴发蓝藻水华,且发生面积、频率和持续时间均呈快速增长趋势(Huismanetal,2018)。水华发生不仅影响感官体验,还可释放多种藻毒素、破坏水生生物网、降低水质、危害渔业生产、社会经济和人类健康(Jeffetal,2019),已成为当今水生态系统...