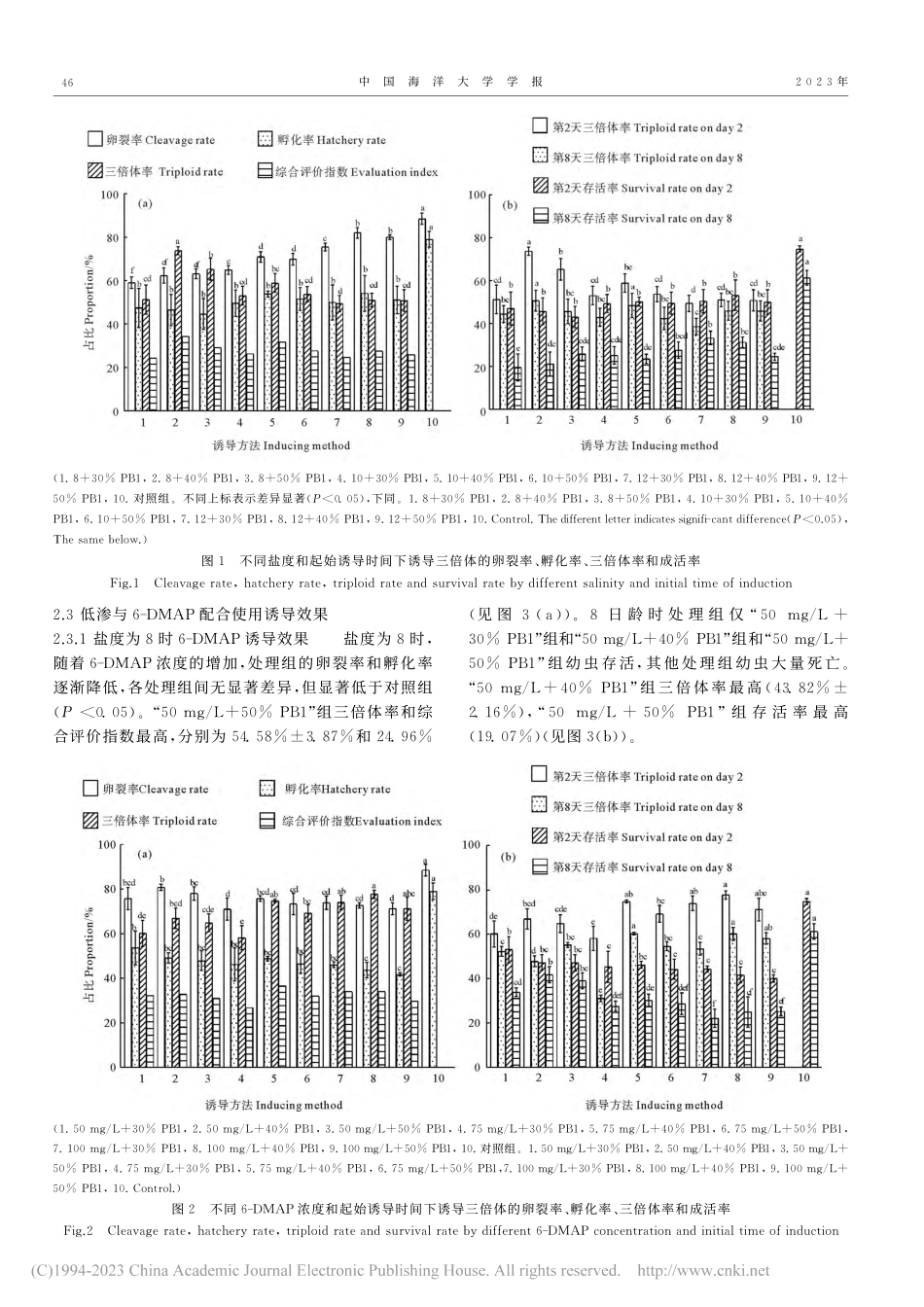
第53卷第2期2023年2月中国海洋大学学报PERIODICALOFOCEANUNIVERSITYOFCHINA53(2):044~050Feb.,20236-DMAP和低渗诱导长牡蛎“海大2号”三倍体的研究*程庚1,徐成勋1,李永国1,李琪1,2**(1.海水养殖教育部重点实验室(中国海洋大学),山东青岛266003;2.青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室,山东青岛266237)摘要:为探索低渗和6-二甲基氨基嘌呤(6-DMAP)诱导长牡蛎(Crassostreagigas)三倍体最佳条件以及6-DMAP和低渗配合使用诱导三倍体的可行性,本研究通过低渗、6-DMAP以及6-DMAP与低渗配合三种方法诱导长牡蛎“海大2号”三倍体。研究显示,低渗处理组中,当第一极体(PB1)出现40%时,在盐度为8的低渗海水中持续处理15min,综合评价指数最高,为34.33%,此时2日龄长牡蛎的三倍体率和存活率分别为73.77%±1.89%和45.78%±6.23%,8日龄长牡蛎的三倍体率和存活率分别为50.65%±4.89%和21.17%±5.54%。6-DMAP处理组中,当PB1出现40%时,75mg/L的6-DMAP持续处理15min,综合评价指数最高(36.52%),此时2日龄长牡蛎的三倍体率和存活率分别为74.81%±0.68%和46.30%±1.54%,8日龄长牡蛎的三倍体率和存活率分别为60.37%±0.61%和30.25%±2.85%。6-DMAP和低渗配合组中,当PB1出现40%时,在盐度为8的低渗海水中,以50mg/L的6-DMAP持续处理15min,综合评价指数最高(36.18%),此时2日龄长牡蛎的三倍体率和存活率分别为74.34%±3.49%和45.07%±4.41%,8日龄长牡蛎的三倍体率和存活率分别为68.23%±2.52%和40.42%±4.30%。综合比较以上三种方法,通过6-DMAP和低渗配合诱导获得三倍体的倍率更稳定,幼虫存活率更高。研究结果可为长牡蛎“海大2号”三倍体培育提供参考资料。关键词:长牡蛎;“海大2号”;三倍体;低渗;6-二甲基氨基嘌呤中图法分类号:S968.3文献标志码:A文章编号:1672-5174(2023)02-044-07DOI:10.16441/j.cnki.hdxb.20220085引...