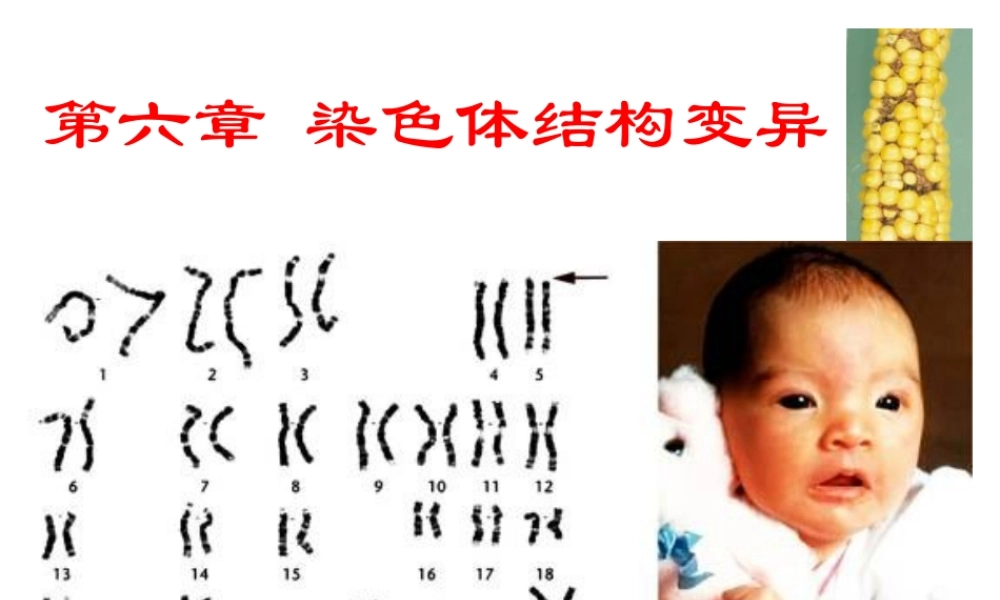
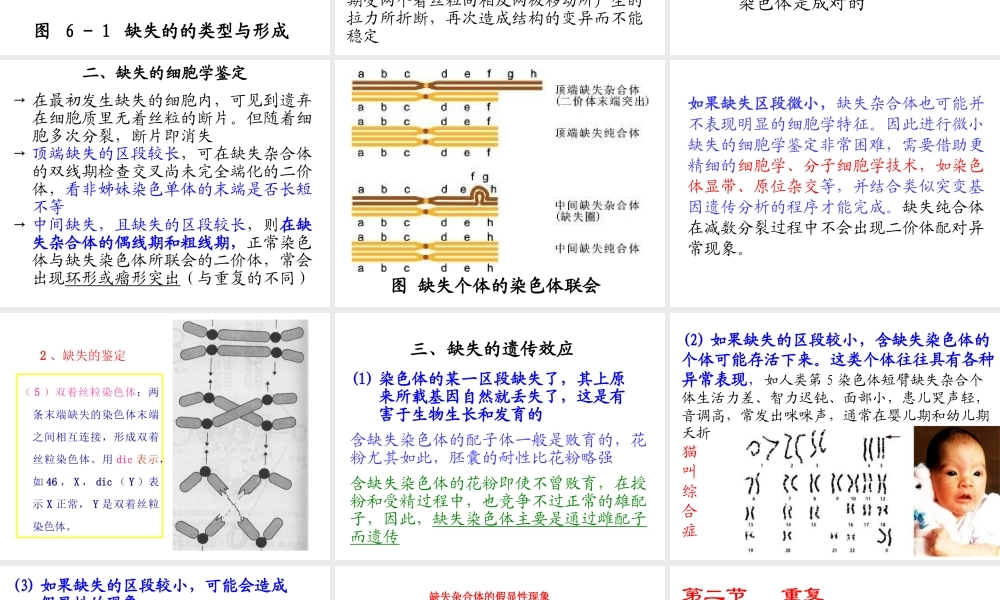
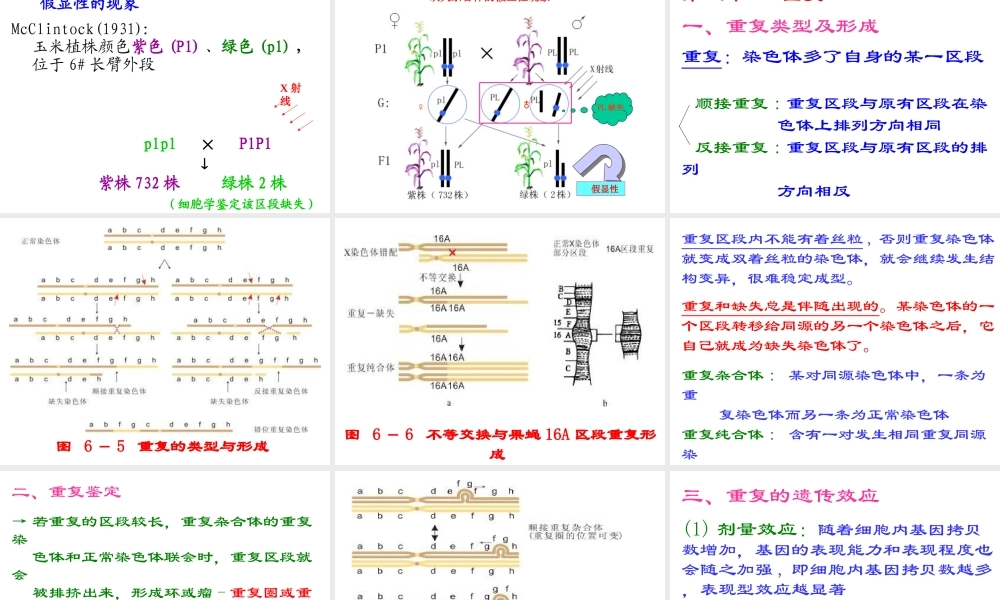
第六章染色体结构变异在自然和人为条件下,可能使染色体折断,之后再接起来,再接合时发生差错,导致染色体结构变异。这种通过“折断—重接”出现的染色体结构变异分为四类:(1)缺失(2)重复(3)倒位(4)易位第一节缺失一、缺失类型及形成缺失:染色体的某一区段丢失了断片:缺失的区段无着丝粒顶端缺失:缺失的区段为某臂的外端某一整臂缺失了就成为顶端着丝点染色体中间缺失:缺失的区段为某臂的内段图6-1缺失的的类型与形成顶端缺失染色体很难定型,因而较少见(1)断头很难愈合,断头可能同另一有着丝粒的染色体的断头重接,成为双着粒染色体(2)顶端缺失染色体的两个姊妹染色单体可能在断头上彼此接合,形成双着丝粒染色体双着丝粒染色体就会在细胞分裂的后期受两个着丝粒向相反两极移动所产生的拉力所折断,再次造成结构的变异而不能稳定中间缺失染色体没有断头外露,比较稳定,因而常见的缺失染色体多是中间缺失的(1)缺失杂合体:某个体的体细胞内杂合有正常染色体及其缺失染色体(2)缺失纯合体:某个体的缺失染色体是成对的二、缺失的细胞学鉴定→在最初发生缺失的细胞内,可见到遗弃在细胞质里无着丝粒的断片。但随着细胞多次分裂,断片即消失→顶端缺失的区段较长,可在缺失杂合体的双线期检查交叉尚未完全端化的二价体,看非姊妹染色单体的末端是否长短不等→中间缺失,且缺失的区段较长,则在缺失杂合体的偶线期和粗线期,正常染色体与缺失染色体所联会的二价体,常会出现环形或瘤形突出(与重复的不同)图缺失个体的染色体联会如果缺失区段微小,缺失杂合体也可能并不表现明显的细胞学特征。因此进行微小缺失的细胞学鉴定非常困难,需要借助更精细的细胞学、分子细胞学技术,如染色体显带、原位杂交等,并结合类似突变基因遗传分析的程序才能完成。缺失纯合体在减数分裂过程中不会出现二价体配对异常现象。2、缺失的鉴定(5)双着丝粒染色体:两条末端缺失的染色体末端之间相互连接,形成双着丝粒染色体。用dic表示,如46,X,dic(Y)表示X正常,Y是双着丝粒染色体。三、缺失的遗传效应(1)染色体的某一区段缺失了,其上原来所载基因自然就丢失了,这是有害于生物生长和发育的含缺失染色体的配子体一般是败育的,花粉尤其如此,胚囊的耐性比花粉略强含缺失染色体的花粉即使不曾败育,在授粉和受精过程中,也竞争不过正常的雄配子,因此,缺失染色体主要是通过雌配子而遗传(2)如果缺失的区段较小,含缺失染色体的个体可能存活下来。这...